Bijlage 17: Van Uytvanck J., 2020, Bosontwikkeling op begraasde voormalige landbouwgronden in het Bos t’Ename, Rapport INBO
Waarom begrazen voor bosuitbreiding?
Sinds de jaren tachtig worden in een hele reeks natuurgebieden in Europa grote grazers zoals runderen, paarden, ezels en schapen ingezet bij het natuurbeheer (WallisDeVries et al., 1998). Waar dit vroeger vnl. gebeurde om het oude cultuurlandschap en de daaraan verbonden specifieke natuurtypes te beheren (patroonbeheer), wordt er de laatste jaren steeds meer gekozen om via begrazing natuurlijke processen te herstellen (als deel van procesbeheer) (Hodder et al. 2005; Holl & Smith 2007). Vaak bestaan grote delen van beheerde natuurgebieden uit voormalige landbouwgronden, die intensief gebruikt en bemest werden. Dit is ook het geval in het Bos t’ Ename waar sinds 2004 ± 26 ha grasland en 11 ha akker samen met 26 ha bos in een groot te begrazen blok werden ingerasterd. Het centrale idee is dat grote grazers de omvorming naar een meer natuurlijk ogend én functionerend landschap kunnen sturen of begeleiden (Bokdam & Gleichman 2000). Daarbij horen ook diverse boslandschappen, hoewel er nog weinig geweten is over hoe zo’n begraasd bos er zou uitzien op langere termijn.
In het Bos t’ Ename hadden de beheerders uiteraard ook gewoon de graslanden en akkers actief kunnen bebossen, zoals de mens dit al eeuwen doet in grote delen van Europa. Uiteraard zal deze optie steeds belangrijk blijven op Vlaamse schaal, maar daarnaast groeit ook het inzicht dat er ook ruimte moet zijn voor meer spontane bosvorming en bosvorming die gestuurd wordt door extensieve begrazing. Begrazing van bossen, heides, bosweides en wastines (woeste gronden) kent overigens een eeuwenoude geschiedenis in Europa (Tack et al. 1993). De keuze voor begrazing gebeurt meestal wel overwogen: de beheerder verwacht dat door het graasgedrag van grote herbivoren, zoals runderen en paarden, de bosvorming veel geleidelijker (en in zekere zin ook natuurlijker) zal verlopen, en dit zowel in de ruimte als in de tijd. Hiermee bedoelen we dat de bosvorming op het terrein via een grillig en vleksgewijs patroon zal verlopen en dat dit patroon zich ontwikkelt over een lange periode, en niet overal tegelijkertijd. Hierdoor ontstaan vele overgangsvegetaties in een mozaïek van kort en lang grasland, ruigtes, struikjes en struwelen, individuele bomen, bomengroepjes en bos met een gevarieerde leeftijdsstructuur. Deze mozaïeken veranderen van vorm, aard en ligging naarmate de tijd verstrijkt en worden daarom ook wel schuivende mozaïeken genoemd (Olff et al. 1999). Dergelijke mozaïeken van verschillende successiestadia naast elkaar in de ruimte worden vaak geassocieerd met een rijke flora en fauna (van Wieren 1998; Dennis et al. 1998). In contrast daarmee ontstaan er bij actieve bebossing scherpere grenzen, is de verticale structuur veel homogener en is ook het bomenbestand nagenoeg gelijkjarig. De spontaan ontwikkelende bossen (zonder begrazing door grote grazers) zitten hier qua structuur ergens tussenin. Zeker in de beginfase van nieuwe bossen (± eerste 30 jaar) zijn er zeer grote verschillen in structuurrijkdom tussen spontaan (hogere structuurrijkdom) en aangeplant bos (Verstraeten et al. 2006). Vele soorten dagvlinders bv. verkiezen juist structuurrijke biotopen met geleidelijke overgangen (zoals boszomen) en het is dan ook niet verwonderlijk dat er juist in deze groep veel bedreigde of zeldzame soorten voorkomen (Maes & Van Dyck 1996).
De effecten van procesbeheer met een grote mate van spontane ontwikkeling zijn echter steeds complex. Wat begrazing betreft, wordt er uitgegaan van het idee dat de effecten van wilde herbivoren voor een deel kunnen worden nagebootst door de introductie van gedomesticeerde dieren. In de praktijk betekent dit echter dat de structuur van het landschap en de soorten die er in leven, veel minder controleerbaar en voorspelbaar zijn dan met klassieke beheersmaatregelen (bv. maaien, kappen, planten, snoeien, begrazen op perceelsniveau) het geval is. Men introduceert immers een moeilijk voorspelbare factor, met name de grote herbivoren zelf, met een gedrag en een levenscyclus die afhankelijk kunnen zijn van de soort, het individu, het terreintype, de grootte van het terrein, het aanwezige voedsel... (Cosyns & Hoffmann 2004).
Door onderzoek werd in de voorbij 15 jaar inzicht verworven over de omstandigheden waarin en in welke patronen bosontwikkeling in het begraasde gebied van Bos t’ Ename optreedt. Onder meer de rol van de uitgangssituatie (akker vs. grasland), graasdruk (hoeveel dieren per ha worden ingezet ontwikkelingstijd, aanwezigheid van zaadbomen, vegetatiestructuur (bieden bepaalde vegetatiestructuren bescherming of juist niet?) en tijdelijke onderbrekingen van het graasbeheer werden onderzocht. Na ± 16 jaar begrazing leverde dat opmerkelijke resultaten en nieuwe inzichten in het bosontwikkelingsproces.
Verschillende bosontwikkelingsspatronen op graslanden en akkers
Op de voormalige akkers van Bos t’ Ename kwam bosontwikkeling zeer snel op gang. Op de kale bodems die na de laatste oogst in 2003 ontstonden, vestigden zich het volgende jaar al massaal pionierboomsoorten zoals boswilg en ruwe berk. In de graslanden op de voedselrijke bodems in het gebied, gaat de bosvorming heel wat trager, verloopt met andere soorten en via andere mechanismen. De voornaamste oorzaak van die trage ontwikkeling is dat bomen moeilijk kiemen in een dichte vegetatiemat én omdat jonge boompjes die er toch in slagen te kiemen samen met het lekkere gras errond worden opgegeten door grote (of kleine grazers).
Bosontwikkeling op voormalige akkers
In voormalige akkers is zeer veel variatie mogelijk, hetgeen door het dynamische karakter van dergelijke milieus te verwachten is. Meestal koloniseren boswilgen, waarvan de zaden honderden meters ver kunnen vliegen de terreinen zeer snel. Ook ruwe berk is een goede windverbreider (zij het iets minder ver) die massaal zaad produceert en verspreidt. Door de geringe afstand tot het bos in Bos t’Ename kwamen grote hoeveelheden zaden op de open bodems van de aanpalende voormalige akkers terecht. Vooral de maand juni lijkt cruciaal voor het kiemen van boswilgen en berken. Als de bodem dan voldoende open is, treedt kieming massaal op. De laatste teelt op de akkers en het al dan niet ploegen erna kan de omvang van die kieming sterk beïnvloeden. Braaklegging na vroege teelten zoals gerst en tarwe en het laten staan van de stoppel zorgt ervoor dat in het najaar en/of het volgende voorjaar grassen die zich vnl. vegetatief voortplanten (bv. kweek en Engels raaigras) en akkeronkruiden zoals vogelmuur de kans krijgen om de vegetatie te sluiten. Hierdoor kan de kieming van bomen bemoeiljkt worden en de bosvorming vertraagd worden. Bij late teelten (maïs, aardappelen, bieten) en wanneer laat in het seizoen nog grondbewerking plaats vindt zal de bodem langere tijd open en losser blijven waardoor de kieming en vestiging van pionierboomsoorten in het volgende jaar gemakkelijker zal verlopen.
Boswilgen en berken zijn snelle groeiers en kunnen pionierbossen voor lange tijd ook domineren (Langeveld et al. 2003). In onbegraasde gebieden ontwikkelen zich op voormalige akkers vaak zeer dichte pionierbossen met duizenden staakvormige stammetjes per hectare. In Bos t’ Ename werd de rol van de grote grazers echter snel duidelijk. Vooral in het vroege voorjaar gaan runderen vaak knabbelen aan de voedselrijke knoppen en smakelijke loten van boswilgen en in mindere mate ook van berken. Hun impact op de structuur van het pionierbos is groot. In de zogenaamde exclosures (door prikkeldraad uitgerasterde vakken waar geen grazers kunnen komen) ontwikkelt zich het hoger genoemde, dichte stakenbos. Dit is bv. goed te zien op locatie 1 op de figuur achteraan deze bijlage. In begraasde zones ontstaan door vraat en betreding meerstammige, wijd uitgroeiende bomen, die sterk verschillen in hoogte en in dichtheid. Daar bovenop spelen de overlevingsstrategieën van de aspectbepalende soorten zoals boswilg en ruwe berk een belangrijke rol. Boswilg hanteert de strategie van de “tolerantie”. Wanneer een smakelijke boswilg (of een andere wilgensoort) wordt aangevreten, kan hij zeer snel regenereren door het aanmaken van nieuwe twijgen, zelfs al wordt hij tot op de grond afgevreten. In Bos t’Ename was en is dit nog goed te zien aan de vaak struikvormige en dichtvertakte kleine boswilgen die dicht op elkaar staan. Ruwe berken zijn veel minder tolerant voor begrazing maar hun overleving is eerder gebaseerd op defensiemechanismen. Berken zijn voor grote grazers onsmakelijke bomen en worden daardoor eerder gemeden. In berken zorgen phenolen in de twijgen ervoor dat de planten onsmakelijk en moeilijk te verteren zijn voor grazers. In Bos t’Ename resulteerden deze verschillen in defensiestrategie bij bomen in een opmerkelijk vestigingspatroon waarbij de berken boven de vraatlijn (de hoogte tot waar grazers aan bomen kunnen knabbelen) en boven de struikvormige begraasde boswilgen konden uitgroeien. Door hun selectieve graasgedrag zorgen grazers op deze manier voor een grote verticale variatie in het zich ontwikkelende bos. Dit is bv. goed te zien in locatie 2 op de figuur achteraan deze bijlage.
De horizontale variatie (verschillen in dichtheid en vlekkenpatronen) wordt in deze pionierbossen vooral in de eerste fase van de bosontwikkeling bepaald. Op dat moment zijn jonge bomen, ook de onsmakelijke berken, het kwetsbaarst. Bij herhaaldelijke begrazing zal een deel van de zaailingen niet overleven. Opnieuw zorgt het selectieve graasgedrag en terreingebruik van de grazers voor deze variatie. Veel gebruikte plekken of routes hebben de grootste kans om tot open plekken of plekjes in het pionierbos te evolueren. Daarnaast speelt ook de aanwezigheid en afstand tot zaadbomen in de omgeving een rol (zie verder).
Bosontwikkeling op voormalige graslanden
De bosontwikkeling in begraasde graslanden verloopt heel anders dan in akkers. De vestiging van houtige soorten is er veel trager, gebeurt in veel lagere aantallen, met andere soorten, in een totaal ander patroon en gaat bijna altijd gepaard met het mechanisme van associatieve resistentie. Dit laatste is naast tolerantie en het vermijden van begrazing (zie hoger bij akkers) een derde mechanisme dat bomen toelaat zich te vestigen in de aanwezigheid van grote grazers. In het kort komt dit mechanisme er op neer dat zich onder extensieve begrazing – met een graasdruk die niet alle jaarlijkse productie wegvreet- vegetatiestructuren in het grasland ontwikkelen die bescherming kunnen bieden aan zich vestigende boompjes. Zaailingen van bomen kunnen opgroeien in minder begraasde ruigtes met stekelige (bv. distels) of onsmakelijke en giftige planten (bv. Jacobskruiskruid), maar vooral in zich ontwikkelende doornige of stekelige struwelen. In voormalige landbouwgraslanden zijn dit vaak bramen, maar na verloop van tijd spelen ook meidoorns, sleedoorns en rozen hier en belangrijke rol.
M.a.w. deze planten zorgen voor bescherming van de jonge boompjes en voldoende grote vlekken van deze planten vormen veilige plekjes (safe sites) voor de uitgroei ervan tot boven de vraatlijn. Dit mechanisme wordt in de ecologie facilitatie genoemd. Deze overleving van zich vestigende bomen is echter veeleer uitzondering dan regel, want veruit de meeste zaailingen van bomen worden samen met het lekkere gras waarin ze opgroeien, opgegeten door grazers. Dit heet associatieve smakelijkheid en is tegengesteld aan het mechanisme van facilitatie. Boompjes worden juist opgegeten omdat ze in een plekje met smakelijk gras terecht zijn gekomen.
In terreinen met een korte ontwikkelingstijd (0-10 jaar) treedt bosvorming in grasland vaak op langs bosranden van waaruit zich bramen uitbreiden. Dit is bv. goed te zien op locatie 3 op de figuur achteraan deze bijlage. In graslanden met een middellange ontwikkelingstijd onder extensieve begrazing (10-20 jaar) is de horizontale structuur van de vegetatie vaak een lappendeken van vlekken met ruigte, kort grasland en ook al kleine en soms grotere struwelen. De ontwikkeling van bv. meidoorn- en rozenstruwelen is vaak het gevolg van de activiteit van muizen en woelmuizen die zaden verstoppen in ruigtes en van vogels die zaden uitscheiden nadat ze de vruchten gegeten hebben. In Bos t’Ename is ook deze ontwikkeling al her en der te zien, zij het voorlopig nog vooral met kleine struwelen. Het is juist in de wat grotere struwelen dat zich kansen voordoen voor de vestiging van loofboomsoorten zoals gewone esdoorn, zomereik, zoete kers en Gewone es, maar ook van minder smakelijke soorten zoals zwarte els. Ook hier worden de zaden vaak aangebracht door vogels (bv. door gaaien die eikels verstoppen) en muizen (die er pitten van kersen in hun holletjes onder het struweel hamsteren). De vestiging van bomen en bosontwikkeling gebeurt dus zeker niet lukraak: in begraasde systemen zijn processen van facilitatie cruciaal en ze interageren met het selectieve graasgedrag van grote grazers en de activiteiten van kleine knaagdieren en vogels. In Bos t’ Ename hebben vooral braamstruwelen een grote invloed op de verspreiding van bomen (horizontale structuur) en op de groei van bomen (verticale structuur). In braamstruwelen, maar ook in ruigtes vestigen zich meer bomen dan daarbuiten en in een aantal gevallen is ook uitgroei boven de vraatlijn (± 2m) reeds mogelijk. Ook meidoornstruwelen fungeren hier en daar als safe site, maar het is te verwachten dat hun rol zal toenemen naarmate de ontwikkelingstijd groter wordt. Nederlands onderzoek wijst op het belang van uitbreidende sleedoornstruwelen voor nieuwe bosvorming (Bakker et al. 2004). Het vestigingspatroon van het jonge bos is van groot belang omdat het reeds in grote mate aangeeft hoe de bosvorming verder zal verlopen (Duarte et al. 2006).
Het belang van zaadbomen
Het belang van zaadbomen werd reeds kort vermeld hierboven. De aanwezigheid van zaadbomen in de onmiddellijke omgeving van het terrein is bijna steeds van belang (zij het in veel mindere mate voor boswilg). De zaadbomen bepalen in eerste instantie de (potentiële) diversiteit van de houtige soorten, in veel mindere mate het effectieve vestigingspatroon. In bos t’ Ename komt een zeer grote diversiteit aan loofbomen voor. Belangrijke vaststellingen zijn echter dat heel wat houtige soorten zich niet of nauwelijks vestigen in de begraasde terreinen, hoewel er in de onmiddellijke omgeving zaadbronnen aanwezig zijn (bv. wilde lijsterbes, tweestijlige meidoorn, tamme kastanje, grauwe wilg, haagbeuk, Spaanse aak, winterlinde, hazelaar, Gelderse roos, … Voor een deel is dit te verklaren door de ecologische vereisten van deze soorten (bv. voor de echte bossoorten). Het zijn echter ook allemaal smakelijke soorten, die in een jong stadium gevoelig zijn voor begrazing. De typische soorten die zich wel in een groot aantal vestigden zijn boswilg en ruwe berk in akkers en gewone es, zomereik, schietwilg, grauwe abeel, zwarte els, eenstijlige meidoorn en sleedoorn in grasland. In de meeste gevallen is er geen duidelijk verband met de afstand tot zaadbomen, behalve in terreinen met een korte ontwikkelingstijd, waar initiële bosvorming vanuit de randen op gang komt, het zgn. bosrand-effect (Walker et al. 2000). Hoe groter de afstand, hoe minder individuen zich vestigen. Hoewel zaden van berken en vooral van boswilgen grote afstanden kunnen afleggen via de wind, zal er in grotere terreinen toch een gradiënt ontstaan waarbij de densiteit van de zich vestigende boompjes afneemt samen met de afstand tot de zaadbomen. Dit is goed te zien op locatie 4 op de figuur achteraan deze bijlage waar zich een veel ijler en graziger bos ontwikkelt verder van de oorspronkelijke bosrand. Dit effect wordt versterkt bij minder goed verbreidende bomen zoals elzen. Ook elzen zijn weinig smakelijk en kunnen zich enkel dicht bij bosranden of zaadbomen snel in voormalige akkers vestigen. Dit is ook goed te zien in Bos t’Ename ter hoogte van locatie 5 op de figuur achteraan deze bijlage. Meestal is echter de vegetatiestructuur uiteindelijk bepalend voor de plaats waar, en welke boomsoort zich vestigt in begraasd terrein.
Graasdruk
In terreinen met een hoge graasdruk, zoals deze wordt toegepast in de gangbare landbouw (met 2 of meer grootvee-eenheden per ha per jaar = min. 730 graasdagen/ha/jaar) kan zich geen bos ontwikkelen, noch op voormalige akkers, noch op graslanden. Bij extensieve begrazing – waarbij dus jaarlijkse productie-overschotten zijn in de vegetatie- ontstaan er kansen voor bosontwikkeling. In Bos t’Ename werd aanvankelijk gewerkt met een kleine kudde van 14 runderen (Oost-Vlaams witrood) en twee Konikpaarden. Rond 2010 werd de runderkudde wat kleiner (een tiental runderen) en werden de Konikpaarden vervangen door afgedankte renpaarden. De dieren blijven het hele jaar door op het terrein. Dit resulteert in een gemiddelde graasdruk van 0,20– 0,25 grootvee-eenheden per ha per jaar. Dit is naar de maatstaven van begrazing onder natuurlijke omstandigheden nog steeds vrij hoog (meer dan 125 kg grazer-biomassa per ha per jaar, WallisDevries et al., 1998), maar is gezien de hoge voedselrijkdom en productie in het gebied een geschikte keuze. De graasdruk kan eenvoudiger uitgedrukt worden aan de hand van het aantal graasdagen per ha en per jaar. Een graasdag is gelijk aan de begrazing van één dier op één dag. In Bos t’ Ename varieerde het aantal graasdagen per ha per jaar in de periode van 2001 tot nu dus tussen de 70 en 93.
Deze gemiddelde graasdruk geeft echter maar een eerste indicatie. Zowel de runderen als de paarden gebruiken het terrein selectief. Observaties in het gebied (data Jan Van Uytvanck, 2008) tonen aan dat de runderkudde ± 63% van haar tijd spendeert in grasland, 24,5% in de verbossende akkers en 12,5% in het bos. 88% van de gespendeerde tijd in het bos vond plaats in de winter, vnl. de late winter. Op het terreingebruik van de paarden hebben we minder zicht, maar vast staat dat ze het overgrote deel van de tijd te vinden zijn op de graslanden en veel minder in het bos gaan. De graasdruk in het terrein varieert dus heel sterk naargelang de aard ervan én het seizoen. In tabel 1 vinden we zo de echte graasdruk in de drie grote landschappelijke eenheden van het gebied gedurende de periode 2004-2020.
Tabel 1: graasdruk in de grote landschappelijke eenheden van het gebied
aantal graasdagen/ha/jaar
grasland 106-141
verbossende akker 98-131
bos 20-27
Uit onderzoek in Bos t’ Ename en 18 andere gebieden op voormalige landbouwgebieden in Vlaanderen (op zwaardere leem- en kleigronden) konden we de relatie tussen graasdruk en bosontwikkeling afleiden in akkers en graslanden (Van Uytvanck 2009, figuur 1). Daarbij zijn twee vaststellingen belangrijk: (1) het verband tussen graasdruk en effectieve vestiging van houtige soorten is niet lineair maar onthult duidelijk drempelwaarden waarboven bosontwikkeling onmogelijk is; (2) in graslanden is een veel lagere graasdruk nodig en is facilitatie (door ruigtes en struwelen) onontbeerlijk, maar ook in akkers waar hogere graasdrukken mogelijk zijn helpt facilitatie de bosontwikkeling sterk vooruit.
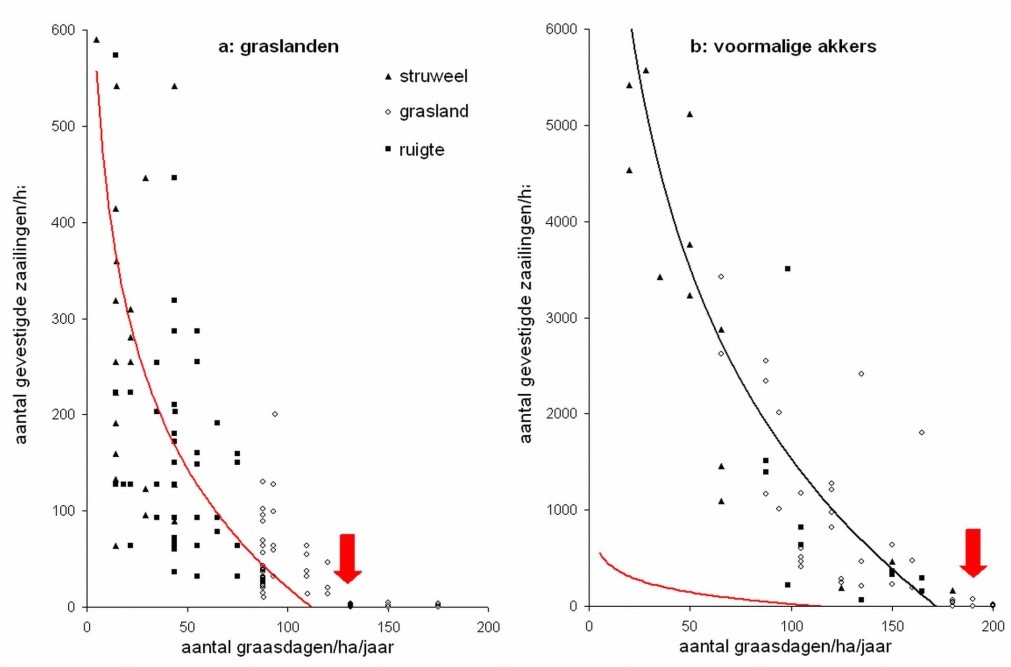
Figuur 1: Vestiging van jonge bomen (> 2m) gedurende de eerste 8-15 jaar na de start van extensief graasbeheer in a. graslanden en b. voormalige akkers. Hoewel elk terrein verschillend is, belemmert een hoge graasdruk spontane bosvorming. De pijlen geven de drempelwaarden aan voor de graasdruk in graslanden en akkers, waarboven geen vestiging optreedt. De rode lijn in (b) is de trendlijn uit (a) om het verschil in schaal te duiden.
Uit figuur 1 blijkt dat de drempelwaarden waaronder bosontwikkeling kan optreden respectievelijk voor graslanden en akkers ± 130 en ±180 graasdagen per ha per jaar bedragen. In akkers is dus een hogere graasdruk mogelijk. Na de vaak massale vestiging neemt het aantal bomen echter meestal terug af. In grasland is een lagere graasdruk vereist, maar neemt het aantal na vestiging meestal toe. Natuurlijk spelen hier ook nog andere factoren mee, zoals de onderlinge verhoudingen van verschillende habitats, kuddestructuur, totale oppervlakte van het gebied enz.. In Bos t’Ename bleek duidelijk dat de graasdruk voor bosontwikkeling onder de drempelwaarde bleef in de voormalige akkers; voor de graslanden lag de graasdruk initieel wellicht te hoog (± 141 graadagen per ha per jaar), maar werd ze na verloop van tijd verlaagd tot ± 106 graasdagen. De verwachting is dat struweelontwikkeling de bosvorming nu op gang zal brengen. Dit is hier en daar al te zien op het terrein. Belangrijk is ook dat de niet verboste delen van de voormalige akkers zich intussen tot grasland hebben ontwikkeld. Hier zal de bosontwikkeling dus ook via de “grasland-manier” moeten verlopen.
Behalve de vastgestelde drempelwaarden is graasdruk dus niet steeds van doorslaggevend belang omdat er selectief gegraasd wordt en er zich nieuwe vegetatiestructuren ontwikkelen die bosontwikkeling kunnen faciliteren. Enkele experimenten in Bos t’Ename en enkele andere gebieden in de regio tonen ook aan dat tijdelijke onderbrekingen van 2 jaar in de begrazing, vooral in de eerste vestigingsfase, de bosontwikkeling fors kunnen vooruit helpen in ruigtes en struwelen die ontstaan in voormalige graslanden (Van Uytvanck et al. 2008). Dergelijke tijdelijke onderbrekingen zijn in de natuur eerder regel dan uitzondering. Grote grazers vertonen van nature trekgedrag, waarbij gebieden tijdelijk onbegraasd blijven. Verder vertonen populaties van grote grazers grote schommelingen, zijn gebieden tijdelijk ontoegankelijk door mechanische barrières of worden ze gemeden door de aanwezigheid van predatoren. In het natuurbeheer is met dergelijke ‘time gaps’ in begrazing tot op vandaag nog te weinig ervaring, maar in theorie is het een valabele optie om meer natuurlijke omstandigheden, zoals het geval is met veel andere beheermaatregelen en zeker bij procesbeheer, na te bootsen.
Hoe zal het bos zich ontwikkelen onder begrazing?
Een ontwikkelingstijd van ±15 jaar zoals die in Bos t’ Ename heeft plaatsgevonden is natuurlijk uiterst kort in vergelijking met de honderden jaren die een volwaardig bos nodig heeft. In het begraasde blok vinden we nu ongeveer 15 ha wastine, waarvan ± 4,5 ha goed ontwikkeld is (met hoge structuurvariatie, zowel op voormalige akker als op grasland). Daarnaast is er aanpalend aan het oorspronkelijke bos 4 ha open pionierbos bijgekomen (op voormalige akkers). Op plekken met spontane bosontwikkeling buiten het begrazingsblok is deze pionierfase al deels achter de rug en nemen secundair gevestigde bomen zoals gewone esdoorn, zomereik en es (voorlopig althans) de overhand op de eerst gevestigde boswilgen. In begraasde terreinen zal dit langer duren en zullen wellicht nog zeer lang open bossen of wastines domineren. Door modelberekeningen voor bosontwikkeling op voedselrijke gronden aan de hand van de gemiddelde groei van bomen, de graasdruk en inzichten verworven in verband met overleving en vestiging van bomen onder begrazing in diverse omstandigheden, is het mogelijk om voorspellingen te doen over hoe een bos er zal uitzien in de komende tientallen jaren (Van Uytvanck 2009). In Bos t’Ename is de verwachting dat onder de huidige graasdruk de kroonsluiting in graslanden langzaam blijft stijgen, maar niet meer dan 20% wordt over een periode van 100 jaar. Voor een volledige kroonsluiting na 100 jaar zou de graasdruk nog 4 keer lager moeten zijn dan nu. De eerste 30-50 jaar is hier een mozaïekvegetatie met vnl. grasland en daarnaast, ruigte en struweelvlekken te verwachten met op beperkte schaal kleine bosschages met o.a. zomereik, hazelaar, haagbeuk, zoete kers en es. Dit is het zogenaamde wastinelandschap. Op de voormalige akkers is de verwachting dat er onder een constante graasdruk zoals nu een volledige kroonsluiting met volwassen bomen optreedt na ± 75 jaar. De eerste 30-50 jaar is hier een open boslandschap te verwachten met vnl. berken, wilgen en elzen afgewisseld met grasland.
De hoger geschetste ontwikkelingen en graasdrukken zijn in de eerste plaats richting gevend. Gebiedseigen, onverwachte en/of nieuwe ontwikkelingen kunnen het bosontwikkelingsproces sterk beïnvloeden (bv. essenziekte, klimaat, gedragsverandering van grazers, nieuwe soorten die opduiken, ...). Maar beheerders hebben dus wel een aantal handvaten om in de tijd en ruimte te spelen met de graasdruk en die aan te passen aan het streefbeeld van het gebied. Bruuske veranderingen zijn echter niet aan te raden omdat de kans op verruiging in een voedselrijk gebied zoals Bos t’Ename groot is (bv. bij forse verlaging van de graasdruk) en omdat er veel kennis dreigt verloren te gaan die zich bij een min of meer constant beheer kan ontwikkelen. Anderzijds is juist het loslaten van bepaalde streefbeelden een keuze die in tijden van klimaatverandering te rechtvaardigen is en die soorten en levensgemeenschappen toelaat zich aan te passen aan die veranderingen.
Besluit
Grote grazers zoals runderen en paarden hebben een grote invloed op de spontane bosontwikkeling op de voormalige landbouwgronden in het gebied. Bosontwikkeling gebeurt in voormalige akkers door pioniersoorten die defensiestrategieën hebben ontwikkeld zoals slechte smaak, gif of graastoleratie. In graslanden hebben bomen de hulp nodig van struwelen (vaak met stekels of doorns) of onsmakelijke ruigtes, waarin jonge bomen (tijdelijk) beschermd zijn tegen vraat. Soms vindt er enkel bosontwikkeling plaats wanneer tijdelijk minder of geen dieren aanwezig zijn. Anderzijds zijn grote grazers juist in staat om bosontwikkeling op gang te brengen. Dit kan door een combinatie van betreding, waardoor microplekjes ontstaan waarin zaden van bomen of struiken kunnen kiemen, en selectief graasgedrag. Afhankelijk van de uitgangssituatie, de aanwezigheid van zaadbomen, de vegetatie in de omgeving en de snelheid waarmee bramen het terrein koloniseren, ontstaan op korte tot middellange termijn open bossen of landschappen die refereren naar de historische bosweidesystemen en wastines. Al deze ontwikkelingen zijn in min of meerdere mate te zien in Bos t’ Ename.
Voor het beheer is het vnl. van belang dat er zich door extensieve begrazing (of ev. door tijdelijke onderbreking in de begrazing) een diverse vegetatiestructuur kan ontwikkelen. In de praktijk hangt veel af van de ontwikkeling van struwelen. Vnl. braamstruwelen brengen bosvorming op gang op voormalige landbouwgronden en structureren er de bosvorming. Geduld, het accepteren van enige onzekerheid en open staan voor verrassingen zijn goede eigenschappen voor de beheerders en bezoekers van het gebied.
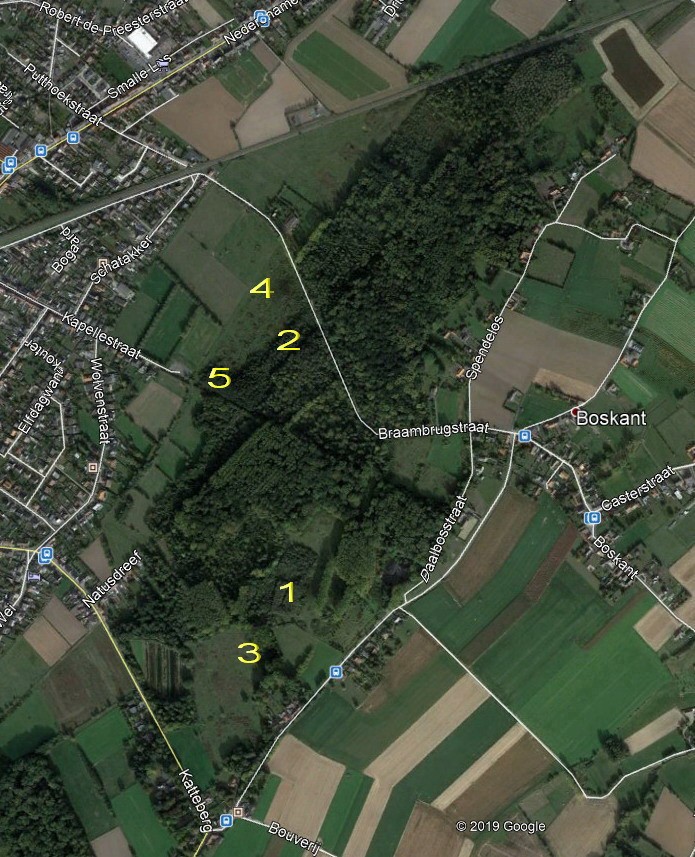
Locaties waar in de tekst naar wordt verwezen
Referenties
Bakker E.S., Olff H., Vandenberghe C., De Maeyer K., Smit R., Gleichman J.M. & Vera F.W.M. 2004. Ecological anachronisms in the recruitment of temperate light-demanding tree species in wooded pastures. Journal of Applied ecology 41, 571-582.
Bokdam J. & Gleichman J.M. 2000. Effects of grazing by free-ranging cattle on vegetation dynamics in a continental north-west European heathland. Journal of Applied ecology 37, 415-431.
Cosyns E. & Hoffmann M. 2004. Extensieve begrazing, mogelijkheden en beperkingen. In Hermy M., De Blust G. & Slootmaekers M. 2004. Natuurbeheer. Davidsfonds Leuven. 362-405
Dennis P., Young M.R. & Gordon I.J. 1998. Distribution and abundance of small insects and arachnids in relation to structural heterogeneity of grazed, indigenous grasslands. Ecological Entomology 23, 23-264.
Duarte L.D.S., Machado R.E., Hartz S.M. & Pillar V.D. 2006. What saplings can tell us about forest expansion over natural grasslands. Journal of Vegetation Science 17, 799-808.
Hodder K.H., Bullock J.M., Buckland B.C. & Kirby K.J. 2005. Large herbivores in the wildwood and in modern naturalistic grazing systems. English Nature, Peterborough.
Holl K. & Smith M. 2007. Scottish upland forests: History lessons for the future. Forest Ecology and Management 249, 45-53.
Langeveld N., de Kort S., Damhuis W. & van Oijen E. 2003. Explosieve bosontwikkeling Meerssenerbroek. Natuurhistorisch maandblad 92, 207209.
Maes D. & Van Dyck H. 1996. Een gedocumenteerde Rode lijst van de dagvlinders van Vlaanderen. Mededeling van het Instituut voor Natuurbehoud.
Olff H., Vera F.W.M., Bokdam J., Bakker E.S., Gleichman J.M., De Maeyer K. & Smit R. 1999. Shifting mosaics in grazed woodlands driven by the alternation of plant facilitation and competition. Plant Biology 1, 127-137.
Tack G., Van Den Bremt P. & Hermy M. 1993. Bossen van Vlaanderen: Een historische ecologie. Davidsfonds, Leuven.
Van Uytvanck J., Maes D., Vandenhaute D. & Hoffmann M., 2008. Restoration of woodpasture on former agricultural land: The importance of safe sites and time gaps before grazing for tree seedlings. Biological Conservation 141, 78-88.
Van Uytvanck J. 2009. The role of large herbivores in woodland regeneration patterns, mechanisms and processes. Doctoraten van het Instituut voor Natuur- en Bosonderzoek 2009 (INBO.T.2009.3). Instituut voor Natuur en Bosonderzoek, Brussel.
van Wieren S.E. 1998. Effects of large herbivores upon the animal community. In: WallisDeVries M.F. (Ed.)., Grazing and conservation management. Kluwer Academic Publishers, Dordrecht, pp. 168-214.
Verstraeten A., Quataert P. & Vandekerkhove K., 2006. Onderzoek naar spontane verbossing en actieve bosaanplant op niet-bosbodems (AMINAL/B&G/38/2002). Deel 1: Vergelijking van de vegetatiestructuur en –soortensamenstelling bij spontane verbossing versus bosaanplanting. INBO.R2006.003
Walker K.J., Sparks T.H., Swetnam R.D., Boatman N.D., Clay D.V., Goodman A., Marrs R.H., Marshall E.J.P., Newman J.R., Putwain P.D. & Pywell R.F. 2000. The colonisation of tree and shrub species within a self-sown woodland: the Monks Wood Wilderness. Aspects of Applied Biology 58, 337-344.
WallisDeVries M. F., Bakker, J. P. & van Wieren S. E. (eds.) 1998. Grazing and conservation management. Kluwer Academic Press, Dordrecht.